所属栏目:推荐论文 时间:2022-05-09

摘 要 以六溴环十二烷(HBCD)为代表的脂环族溴代阻燃剂(CBFRs)被广泛应用于纺织、建材、电子、电气、化工、交通、建材等领域.随着 HBCD 作为《关于持久性有机污染物的斯德哥尔摩公约》增列持久性有机污染物,与 HBCD 具有相似结构和性能的四溴环己烷(TBECH)和四溴环辛烷(TBCO)等 CBFRs 被当作 HBCD 的潜在替代产品.迄今为止,HBCD、TBECH 和 TBCO 已在大气、水体、土壤等多种环境介质和生物体中被检出,它们在生物体内的代谢转化以及内分泌干扰、神经、生殖、发育等毒性效应亦受到广泛关注.值得指出的是,所有 CBFRs 均含有同分异构体,表现出异构体选择性的生物富集、代谢和毒性效应.遗憾的是,目前相关研究还十分匮乏.本文从 CBFRs 的环境暴露水平、生物富集、毒性效应、以及 CBFRs 的生物转化等方面展开综述,特别强调了从异构体水平研究HBCD 及其替代物的必要性.本文有助于全面了解 CBFRs 生物富集、代谢及毒性效应,对于正确认识和准确评价 CBFRs 的生态和健康风险具有重要的科学意义.
关键词 脂环族溴代阻燃剂,异构体,生物富集,毒性效应,生物转化.
溴代阻燃剂是一类包括脂肪族、脂环族、芳香族及芳香-脂肪族的含溴化合物.其中脂环族溴代阻燃剂(cycloaliphatic brominated flame retardants, CBFRs)的阻燃效果好于芳香族溴代阻燃剂,热稳定性能优于脂肪族溴代阻燃剂,而一度作为多溴联苯醚(PBDEs)的替代品被广泛应用于纺织、建材、电子、电气、交通、化工、石油等领域中[1].脂环族溴代阻燃剂主要包括六溴环十二烷(hexabromocyclododecane,HBCD),1,2-二溴-4-(1,2-二溴乙基)-环己烷(tetrabromoethylcyclohexane,TBECH),和 1,2,5,6-四溴环辛烷(1,2,5,6-Tetrabromocyclooctane,TBCO). 目前关于 CBFRs 研究最多的是六溴环十二烷,其作为全球第三大溴系阻燃剂,在生产、使用、运输及废弃物报废处理等过程进入环境.迄今为止,人们已经在许多环境介质、生物体如鸟类[2]、水生动物[3 − 4]、植物[5 − 6]、人体[7],甚至在遥远的北极圈的生物体[8] 内都检测到 HBCD.
随着 HBCD 增列《关于持久性有机污染物的斯德哥尔摩公约》持久性有机污染物,其生产和使用受到限制,人们会选择 TBECH 和 TBCO 这样结构和性质相似、但知名度较低、认识尚不深入的溴代阻燃剂.虽然目前 TBECH、TBCO 的产量远低于 HBCD,但可以预见在不远的未来,这类阻燃剂的产量、使用量将会上升,从而导致其环境水平的增加[9 − 10],因此有必要充分认识这类阻燃剂的环境和生态效应[11 − 16].
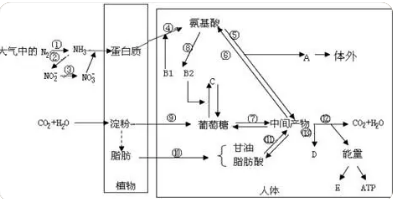
TBECH、TBCO 和 HBCD 一样,具有溴代的脂环结构,包含多种异构体和对映体,但碳环数、溴取代数、取代位置、空间结构存在差异.展示了 HBCD、TBECH、TBCO 的理化性质,其中部分参数来自 EPI Suite 和 SPARC 软件的计算. 相比 HBCD,TBECH 和 TBCO 的分子量较小,熔点较低,稳定性较差;蒸气压和溶解度都较高,说明两者的挥发性和水溶性更大;lgKow 都大于 4,说明三者同样具有较强的亲脂特性和生物蓄积潜力.三者结构和性质的异同对它们的生物富集、代谢及毒性效应的影响尚不清楚. 研究表明,异构体的生物富集、代谢及毒性效应均存在显著差异[13, 17 − 22].只有在异构体水平进行研究,才能全面准确地对 CBRFs 的生态和健康风险做出评价本文对 HBCD、TBECH、TBCO 及其异构体的生物富集、代谢及毒性效应进行综述,并预测了这3 种污染物的研究趋势.
1 CBFRs 异构体的环境暴露水平(Bioaccumulation of CBFRs isomers)
1.1 HBCD 异构体在环境介质中的分布工业级
HBCD 主要含有 α-HBCD、β-HBCD、γ-HBCD 等 3 种异构体,其比例分别为 10%—13%,1%—12% 和 75%—89%,其中 γ-HBCD 是工业品中最主要的异构体,另外工业品 HBCD 中还含有少量的 δ-HBCD 和 ε-HBCD[29].HBCD 可以在从生产到废物处理的整个生命周期中进入环境,展示了HBCD 在各个污染源区环境介质中的水平.天津[30] 聚苯乙烯生产厂附近粉尘、土壤和沉积物中均检测出较高的水平的 HBCD,在中国香港[31] 污水处理厂的污泥中,HBCD 浓度比在悬浮颗粒物中的浓度高出近 2 个数量级,说明其容易在污泥中发生富集,α-HBCD 为主要异构体.HBCD 在华北塑料垃圾处理区[5] 的土壤中的浓度大于处理厂路边和农田土壤中的浓度,γ-HBCD 是土壤中主要的异构体.其比例可能受到 HBCD 加工过程中热异构化和环境介质中非生物/生物转化的影响.同样发现在中国渤海[32] 附近一家 HBCD 生产企业和捷克源区[33] 周围的土壤和沉积物中,HBCD 异构体比例与工业品相似,都以γ-HBCD 为主,推测 HBCD 在土壤等不透光封闭性较强的介质中不易发生异构体转化,此外在捷克源区的产品仓库和工厂外大气、消费产品和建造材料中都检测出了以 γ-HBCD 为主的 HBCD,其比例与工业品相似,可见上述介质中的污染直接来源于 HBCD 工业品的生产和使用。
在捷克源区的房屋室内和室外背景空气中观察到了工厂空气中 95% 为 γ-HBCD,而背景空气中 40% 为 γ-HBCD,表明存在不同异构体的大气迁移和 γ-HBCD 到 α-HBCD 的光转化,工业活动中 HBCD 的大量生产和使用正在造成持续的环境污染.上述研究表明,HBCD 的生产和使用,导致 HBCD 进入大气、水体和土壤等环境介质.进入环境的 HBCD,其异构体比例可能发生变化[30 − 33].HBCD 作为溴代阻燃剂,已被确定为室内和室外环境中的全球污染物. 研究表明,不同国家、地区的大气气相和颗粒相中都可以检测到 HBCD[33 − 38].瑞典斯德哥尔摩[36] 室内灰尘中 HBCD 的平均浓度为 190 ng·g−1.
中国大连大气[37] 中 73%—97% 的 HBCD 存在于颗粒相. 在深圳[38] 室内大气颗粒相中,96% 以上的 HBCD 都存在于 PM10. 无论是气相还是颗粒相,α-HBCD 都是主要组分,推测 α-HBCD 更容易被吸附在灰尘等颗粒物表面.捷克源区[33] 房屋室内和室外背景空气中的 HBCD 都以 α-HBCD 为主,γ-HBCD 的比例明显小于工业品,推测 HBCD 在空气中发生异构体转化,室外空气中的污染可能来自于暖通空调系统、装饰性聚苯乙烯和绝缘材料. 可见,大气颗粒物是 HBCD 的重要归趋,HBCD 的室内浓度高于室外浓度.
此外,据报道来自中国[39] 的空气样本中,δ-HBCD 浓度高达 15%,超过了在商品HBCD 中的比例.但是并不确定 δ-HBCD 的来源.工业废水的排放、大气沉降等过程会造成 HBCD 持续的水体污染.在韩国[40] 工业园区附近的湖泊中,HBCD 在沉积物中的浓度与中国天津市大沽河[41] 和中国贵屿连河[42] 中的浓度相似,但比日本九头龙河[43] 沉积物中的浓度低 1 个数量级,在该湖泊附近的小溪中也检测到了 HBCD. 有研究报道我国沉积物中 HBCD 的浓度从东到西呈现下降趋势. 天津大沽河[41] 中 HBCD 的总浓度最高,其次是天津港[41] 和长江[44]. 大沽河、天津港、天津地区 HBCD 总浓度较高的原因可能是受城市和重工业活动的影响.
在东江和珠江口的沉积物中,HBCD 的最高平均浓度达到 64.7 ng·g−1 dw[45]. 从全国 HBCD 的分布来看,天津、长江和广东的 HBCD 总浓度高于其他地区,这主要是由于这些地区是人口密集区和工业活动 密 集 区 并 对 HBCD 的 市 场 需 求 较 高 . 在 渤 海 南 部 海 域 未 过 滤 水 体 中 , α-HBCD、 β-HBCD 和γ-HBCD 浓 度 范 围 分 别 为 1.23 —1800 ng·L−1、 0.85 —1120 ng·L−1、 1.10 —2150 ng·L−1, α-HBCD 和γ-HBCD 是主要成分[32]. 结合相关研究[32, 43, 46] 发现我国渤海南岸的浓度最高,同时也是世界上浓度最高的,因为我国渤海南岸靠近 HBCD 生产地. 研究发现沉积物中 HBCD 的异构体组成与工业品中的相似,均以 γ-HBCD 为主,而在活性污泥中则以 α-HBCD 为主,其原因是 β-和 γ-HBCD 均可以转化为α-HBCD 以及微生物对 α-HBCD 的降解速率最慢[47].污水处理厂的污物处置[47]、垃圾填埋地的渗漏、塑料及电子垃圾的不当处置[5] 都会导致土壤中HBCD 的污染.
除此之外,土壤中的 HBCD 还可能来自大气中的干湿沉降[33].HBCD 由于其疏水性,可以与固体颗粒如土壤、沉积物和污泥有很强的结合. 研究发现不同类型土壤之间也存在显著差异.Tang 等[48] 分析了浙江省宁波市的垃圾填埋场、工业区、居住区、交通区、菜地和农田土壤,发现在废物倾卸场地和工业区的 HBCD 浓度较高,HBCD 在菜地和农田土壤中的污染极有可能是由扩散源引起的. 与潍坊和宁波相比,除了广东清远,广东的工业区和电子垃圾回收站 HBCD 浓度最高[49],与印度、越南、马来西亚、印度尼西亚和柬埔寨的露天倾倒场土壤的平均值(<2.4 ng·g−1 dw)相当[50]. 华南地区 HBCD 浓度较低的原因可能与采样点有关.Li 等[51] 报道从制造设施开始 HBCD 浓度呈下降趋势.山东、浙江和广东土壤中 HBCD 异构体分布相似,其中 γ-HBCD 为主要成分,α-HBCD 和 β-HBCD 次之[48 − 49, 52].土壤中 HBCD 的异构体组成与工业品中的相似,均以 γ-HBCD 为主[47].展示了 HBCD 在不同地区大气、水、土壤等介质中的环境水平.
1.2 TBECH 和 TBCO 异构体在环境介质中的分布
TBECH 有 4 种异构体,分别是 α-TBECH、β-TBECH、γ-TBECH、δ-TBECH,其中商业品(t-TBECH)中以 α-TBECH 和 β-TBECH 为主要组分,二者比例约为 1∶1[26],而 β-TBECH 是环境中较为常见的异构体[53, 54].商业品 TBCO 有 2 种异构体:α-TBCO、β-TBCO,其比例分别为 34% 和 66%[55].与大量关于 HBCD 的报道相比,环境中 TBECH 和 TBCO 的报道非常有限,而与 TBECH 相比,TBCO 的报道更是少之又少. 作为新型溴代阻燃剂,TBECH 和 TBCO 在建筑材料、电子设备、家具用品中的使用,使其很容易挥发到室内空气中,进而污染室外空气. 如今已在挪威和英国[56] 的室内大气和灰尘中检测到了 TBECH,办公室布置相比家庭,具有较快的器具及电子产品更换速率,使得办公室大气中 TBECH 浓度普遍高于家庭房屋中的浓度,α-TBECH 为主要异构体。
室内灰尘中 TBECH 的浓度相比大气要高得多,在家庭和办公室室内灰尘中,β-TBECH 为主要异构体,可能是由于 β-TBECH 相比 α-TBECH 具有较低的挥发性,更容易在灰尘中积累.意大利罗马[57] 家庭和工作场所的室内灰尘样本(33.7—228 ng·g−1),美国加利福尼亚州[58](<0.64—360 ng·g−1;54% 的检出率)在室内环境中测得的TBECH 平均浓度是迄今为止最高的. TBECH 在瑞典斯德哥尔摩[36] 室内和室外大气、室内灰尘中也有所检出,公寓中的浓度是办公室浓度的 4 倍,这与上述英国家庭和办公室内浓度的结论不同,说明TBECH 的浓度可能很大程度上受到室内装修布置的影响.
同样在捷克[59] 发现不同房屋大气中TBECH 的浓度差距很大,室内最高浓度比第二高浓度高出 10 倍;其夏季浓度高于冬季,夏季和冬季的室外平均浓度分别为 96、2.2 pg·m−3,体现出了在夏季 TBECH 更高的挥发性.在英国西米德兰兹[34] 也发现 夏 季 TBECH 的 浓 度 高 于 冬 季 , 小 于 英 国 [56] 室 内 空 气 浓 度 的 报 道 , 其 检 出 量 是 瑞 典 空 气 的10—20 倍.在瑞典新建幼儿园[60] 空气和灰尘、中国北方电子垃圾回收区大气[61] 中检测到 TBECH 和TBCO 异构体,后者以 β-TBECH 和 β-TBCO 为主要组分,TBCO 在后者中的浓度为 30 pg·m−3. 有报道发现 TBECH 具有持久性[62] 和长距离迁移性[63],在偏远的北极和南极地区已经测量到 TBECH 的存在. 在北 极 和 亚 北 极 [64] 检 测 到 了 TBECH 的 大 气 沉 积 .
在 来 自 挪 威 北 极 地 区 [65] 的 被 动 空 气 样 本 中 ,α-TBECH 和 β-TBECH 是检出率最高的卤化阻燃剂,每种异构体的浓度分别为 29—65、17—46 pg·m−3.虽然无法获得北极其他地区(例如北美)空气中 TBECH 的浓度,但在加拿大北极地区的一些海洋生物群中检测到了 β-TBECH,表明其具有远距离运输的潜力[66]. 最近一个报告指出,在南极洲西部的南设得兰群岛大气样本中检测出 α-TBECH、β-TBECH 和 β-TBCO,主要分布在颗粒相中,浓度范围分别为nd—0.39、nd—1.3、nd—0.21 pg·m−3.这意味着 TBCO 可能也具有环境持久性和长距离迁移能力[9].通过污染源的排放和大气沉积,TBECH 和 TBCO 会进入地表水、沉积物、底泥和土壤.如今已在中国北方电子垃圾回收区[61] 的水、土壤中检测出 TBECH,且以 β-TBECH 为主;TBCO 在土壤和灰尘中的浓度分别为<0.0493—0.0890 ng·g−1dw、7.38 ng·g−1,以 β-TBCO 为主.
中国香港水域[67] 地表沉积物中的 TBECH 浓度比该水域中 HBCD 的浓度低约 1 个数量级,且发现两者浓度存在很强的相关性,推测两者的排放源或排放途径相似.中国香港污水处理厂[31] 中废水(包括颗粒相和溶解相)和污泥中TBECH 的浓度比该研究中 HBCD 相应浓度低一个数量级,底泥中浓度高出悬浮颗粒物两个数量级,且都以α-TBECH 为主,意味着 TBECH 的污染和扩散程度小于 HBCD. TBECH 与 HBCD 具有相似的理化性质,与固体颗粒如污泥、沉积物有很强的结合.新加坡[68] 地表水和悬浮沉积物中 TBECH 的浓度与中国香港污水处理厂的报道不同,底泥沉积物中的浓度低于悬浮沉积物,可见 TBECH 在地表水体中并未发生大量沉积.在德国北海地区[69] 的沉积物中检测到高达 12 ng·g−1dw 的 TBCO,这意味着 TBCO 从河流到北海沉积区的迁移率很高.可见,作为 HBCD 的替代品,TBECH 和 TBCO 已经在大气、水体、土壤等多种环境介质中被检出.展示了 TBECH 在不同地区大气、水、土壤等介质中的环境水平.
2 CBFRs 的生物积累(Bioaccumulation of CBFRs)
2.1 HBCD 在生物体中的积累和分布生物体
能通过环境暴露和食物链传递蓄积 HBCD. 一系列调查结果表明,HBCD 在水生和陆地生态系统中都存在生物富集现象.有关海洋生态系统的调查发现,在 2005 年至 2015 年之间,中国南海江豚 (Neophocaena phocaenoides)和 海豚 (Sousa chinensis)[70] 中 HBCD 的浓度范围分别为 97.2—6260 ng·g−1lw 和 447—45800 ng·g−1lw(lipid weight,以脂质重计),明显高于其生长环境,且 HBCD 主要积累在脂肪中,含量逐年增长.
在渤海[71] 生态系统所有生物样品中 HBCD 的浓度范围在 3.45—461 ng·g−1dw 之间,在污染最重的鲻鱼(Liza haematocheila) 中高达 332 ng·g−1dw. 中国香港水域[67] 海洋食物网中的生物体中 HBCD 的浓度范围在 3.01—93.2 ng·g−1lw 之间,生物放大因子(BMF,捕食者和猎物之间平均脂质标准化浓度的比率)都大于 1,意味着 HBCD 发生了生物富集和食物链放大,该现象同样在渤海[71] 沿岸和南极洲乔治王岛[72]的生态系统中被发现. 在乔治王岛生态系统的所有样品中均检测到 HBCD,α-HBCD 和 γ-HBCD 在所有样品中占主导地位,α-HBCD 的 TMF(营养级放大因子,描述平均脂质归一化浓度与营养级之间的相关性)显著大于 1.
上述调查结果证明了 HBCD 在海洋生物中普遍存在生物富集和食物链放大现象. 目前认为 α-HBCD 的富集能力最强[67, 70 − 71],有报道称 γ-和 β-HBCD 不具有生物富集能力[71].有关淡水生生态系统的调查发现,在韩国河流的鰕虎鱼 (Gobiidae)[47] 中,HBCD 的浓度范围在24.96—66.71 ng·g−1lw 之间. 韩国工业园区鲫鱼 (Carassius carassius)[73] 不同组织 HBCD 的积累量为:肝脏>卵>血液>肌肉,不同组织中 HBCD 的浓度普遍高于水体和沉积物中的浓度,所有组织中的生物富集因子(BCF)都大于 1,意味着在鲫鱼中发生了生物积累. 韩国工业园区附近的湖泊[40] 中,HBCD 在鱼类和贝类中的平均浓度分别为 1.55 ng·g−1ww(wet weight,以湿重计)、60.9 ng·g−1lw,且 α-HBCD 相比γ-HBCD 在鱼类和贝类中更占优势。
其生物群沉积物累积因子(BSAF,生物体内的浓度经过脂质归一化,沉积物中的浓度经过 TOC 归一化)在 0.79—3.21,此外还观察到近岸的生物样本(平均值:2.42)中BSAF 高于远离海岸的生物样本(平均值:1.73). 在华南某电子垃圾场的捕食性鱼类鲮鱼 (Cirrhinusmolitorella) 和北方蛇头鱼(Ophicephalus argus)[74] 中,HBCD 的浓度分别为0.07—96.9、0.18—240 ng·g−1ww,BCF 分别高达 6225 和 6431.
不同组织 HBCD 的积累量为:脂肪>肝脏>鳃>肌肉,积累量与组织脂质含量正相关,说明 HBCD 更容易蓄积在生物脂肪中. 在电子拆解区水域[75] 中,苹果螺 (Ampullaria gigasspix) 和草鱼 (Ctenopharyngodon idellus) 体内 HBCD 的平均浓度分别为 5.02 ng·g−1lw 和 45.9 ng·g−1lw,HBCD 在水生食物网中的 TMF 为 6.36,表明存在生物放大. 上述调查结果说明 HBCD 在淡水生物中也普遍存在生物富集现象,浓度水平与在海洋生物中的相同. 研究发现,在鲤鱼 (Cyprimus carpio morphamoblis)[76] 中存在 α-和 γ-HBCD 的特异性富集,α-HBCD 在不同组织中的 BCF 均显著高于 β-和 γ-HBCD.在天津渤海的海洋生物中,鲱鸥(Larus argentatus)体内 HBCD 的浓度最高(1111 ng·g−1 lw),在渤海的海洋和淡水食物网中 α-HBCD 的浓度和 HBCD 的总浓度随着营养水平的增加而增加,说明在所有异构体中 α-HBCD 的生物富集潜力较强[77].与大量的 HBCD 在水生生物中的富集报道相比,HBCD 在陆生生态系统中生物积累研究相对较少,主要集中于植物[5, 78].
研究者在户外松针(Pinus armandi Franch, 208 ng·g−1dw)[79]、南极的苔藓(Andreaea depressinervis、Sanionia uncinate, 0.63—930 pg·g−1dw) 和 地衣 (Himantormia lugubris、Usneaantarctica, 0.1—21.1 pg·g−1dw)[80] 中均检测出 HBCD,说明植物能够吸收空气中的 HBCD,同时也意味着 HBCD 能够进行远距离大气传输. Zhu 等[81 − 82] 的研究也证明植物既可以通过叶片吸收空气中的HBCD,也可以通过植物根吸收土壤中的 HBCD,根吸收的 HBCD 能够传输到地上部分. 在珠江三角洲各类蔬菜[83]、自然生态系统红树林[78] 中观察到 HBCD 在植物组织中分布为茎>根>叶.
无论是水生还是陆生生物,对 HBCD 的吸收均显示出显著的异构体差异性.相比 β- HBCD、γ-HBCD,α-HBCD 具有较高的水溶性[78]、TMF[71]、BCF[73]、生物积累因子(BAF)和摄取速率常数[84],较长 的 半 衰 期 [84 − 86], 较 低 的 降 解 率 [87], 且 β- HBCD、 γ-HBCD 会 在 生 物 体 内 异 构 化 [76, 84, 88 − 89]为 α-HBCD,因此,α-HBCD 在生物体内中更占优势. 大量研究发现,HBCD 异构体在生物体内的浓度大小 为 α-HBCD>γ-HBCD>β-HBCD, 该 规 律 被 发 现 存 在 于 草 鱼 (Ctenopharyngodon idellus)[75]、 斑 马 鱼(Brachydanio rerio var)[85]、蚯蚓 (Eisenia fetida)[90]、鸭 (Anas platyrhynchos)[86]、牛 (Bos taurus)[91]、加拿大顶级捕食性鱼类 (Salvelinus namaycush、Stizostedion vitreum、Salvelinus fontinalis)[92]、日本虎斑猛水蚤(Tigriopus japonicas)[93]、捕食性鱼类鲮鱼和北方蛇头鱼[74] 以及渤海生态系统和韩国河流的鱼类[47, 71]中 .
也 有 不 符 合 上 述 积 累 顺 序 的 , 如 在 渤 海 的 虾 (Fenneropenaeus chinensis) 和 蛤 蜊 (Ruditapesphilippinarum) 中,HBCD 异构体浓度大小 γ-HBCD>α-HBCD>β-HBCD,这可能是由于虾和蛤蜊位于食物链底层,直接受到其环境中异构体比例的影响[71]. 陆生植物的研究显示,大部分植物[5, 78 − 82] 中α-HBCD 为主要异构体,但在珠江三角洲蔬菜[83] 中,γ-HBCD 为主要异构体,α-HBCD 的比重与土壤接近,蔬菜根部对 HBCD 不具有异构体选择性吸收. 有学者发现,在沿渤海生态系统中,3 种 HBCD 异构体在不同生物中的分布比例存在差异[71]. 沿着虾、蛤蜊、鰕虎鱼 (Acanthogobius flavimanus)、鲈鱼 (Lateolabrax japonicus) 这一食物链,随营养级的增加,α-HBCD 的比例增大,γ-HBCD 的比例降低,由此推测出 α-HBCD 具有生物放大作用.
2.2 TBECH、TBCO 在生物体中的积累和分布
TBECH 和 TBCO 可通过饮食摄入、皮肤接触或呼吸吸入等途径进入生物体内.目前,国内外关于二者在环境生物中积累和分布的研究主要集中于水生生态系统. 一项关于太湖食物网[94] 的研究发现,β-TBECH 在 10 种水生生物中的浓度范围为(276±628) pg·g−1ww,而 α-TBECH 仅在银鱼 (Protosalanxhyalocranius) 中被检测到((134±208)pg·g−1ww). 迄今为止,已在鲱鸥(Larus argentatus)[53, 95 − 96] 中持续检测到 TBECH 的存在. Ruan[70] 的研究发现,中国南海海豚脂中的 TBECH 从 2005 年到 2015 年的十年间 显 著 增 长 , 江 豚 (Neophocaena phocaenoides) 和 海 豚 (Sousa chinensis) 中 TBECH 的 浓 度 范 围 为<0.8—125、<0.8—362 ng·g−1lw,其中 α-TBECH 为主要异构体,其浓度比该研究中 HBCD 的浓度低约2 个数量级。
但高于环境中浓度. 中国香港附近水域[67] 的海洋食物网中,TBECH 的浓度范围在<0.8—13.9 ng·g−1lw,α-TBECH 为主要异构体,在软体动物中的浓度明显高于甲壳类和鱼类中的浓度,表明不同生物对 TBECH 的积累存在差异.但是 TBECH 的 BMF 都大于 1,与 HBCD 相比,营养级放大因子较低,说明 TBECH 具有生物放大潜力但低于 HBCD.TBECH 在甲壳和软体动物中的 BSAF 均小于 1,意味着 TBECH 在底泥生物中并未发生生物富集. 对于不同异构体,γ-TBECH 和 δ-TBECH 的BMF 显著高于 α-TBECH 和 β-TBECH,即 γ-TBECH 和 δ-TBECH 的生物放大潜力相对较高,但一般环境中以 α-TBECH 和 β-TBECH 为主要组分,因此生物体中也以 α-TBECH 和 β-TBECH 为主要组分. 研究表明,在鲸类动物体中,α-TBECH 占主导地位.最近的调查结果显示,食物样品中 β-TBECH 浓度远大于英国室内空气和尘埃。
在野生鲭鱼 (Scomberomorus niphonius) 和金枪鱼 (Thunnini) 中发现 TBECH 的浓度分别为 6.0、39.48 ng·g−1lw,β-TBECH 是主要异构体;在食物和母乳[97] 中出现频率最多、含量最高的 EFR(新型阻燃剂) 是 β-TBECH,浓度为 2.5 ng·g−1 lw,与 α-HBCD 的浓度 (2.1 ng·g−1 lw) 相当.TBCO 在水生生物体内亦有发现.在北美五大湖的水鸟银鸥 (Larus argentatus) 蛋[53] 中检测出TBCO. 在德国湾的黄盖鲽 (Limanda limanda)[69] 中检测到 TBCO,其最高浓度为 12 ng·g−1 ww. 在安大略湖的鳟鱼 (Salvelinus namaycush) 和圆虾虎鱼 (Neogobius melanostomus)[98] 中检测到 TBCO 浓度分别为0.033、0.009 ng·g−1ww. 2003 年欧洲环境署科学小组的研究报告指出,TBCO 符合欧盟的 vPvB 筛选标准,即具有显著的持久性和生物累积性[99].研究发现 TBCO 在海水中及淡水中的半衰期大于 60 d,在海洋及淡水沉积物中半衰期大于 180 d.
然而目前对 TBCO 生物积累的认识还非常缺乏. 在日本青鳉(Oryzias latipes)[16] 胚胎试验中发现 TBCO 在胚胎中的累积速率常数为 1.7—1.8 d−1,生物富集因子最高达到 1.3×104,显著大于 HBCD 意味着其具有较强的生物富集能力,从而影响胚胎发育. 在德国北海地区,TBCO 在底栖鱼类[69] 中浓度最高为 12 ng·g−1ww,其 BSAF 为 2.9,接近于已知的持久性生物有机污染物 BDE-47(2,2',4,4'-四溴联苯醚)和 BDE-100(2,2',4,4',6-五溴联苯醚)的 BSAF,表明该研究中底栖鱼类对 TBCO 具有生物富集作用. 但 TBCO 异构体水平的生物积累知之甚少,植物作为环境中重要的生物介质,对有机污染物的环境行为有着不可忽视的作用.目前关于植物中 TBECH、TBCO 的吸收、传输和转化过程知之甚少.
3 CBFRs 异构体的生物代谢及转化(Biological metabolism and transformation of CBFRs isomers)
3.1 HBCD 的生物转化
3.1.1 HBCD 的生物异构化异构化是改变化合物的结构而分子量不变的过程.HBCD 的立体结构复杂,主要包含 3 种立体异构体,分别为 α- HBCD、β- HBCD 和 γ-HBCD. 不同 HBCD 异构体对生物产生的毒性效应存在差异,因此了解不同异构体的生物异构化现象十分必要. 早期的研究显示,HBCD 在生物中不存在异构体转化现象.如 Essslinger 等[100] 用含纯 γ-HBCD 的食物喂养镜鲤(Cyprinus carpio morpha noblis),并未观察到γ-HBCD 异构化为 α-HBCD 的证据. 随着研究的深入,有关 HBCD 异构体在不同生物体内的异构化的报 道 不 断 涌 现.
Gannon 等[101] 用 含有 98% γ-HBCD 的 商品 HBCD 喂 养 大鼠 (Fischer F344、 SpragueDawley、Wistar),在中、高剂量投喂下的大鼠组织中均检测到 10% 左右的 α-HBCD,证明了 γ-HBCD 向α-HBCD 转化的生物异构化. 与 γ-HBCD 相比,α-HBCD 更易分布在肝脏.在母鸡(Gallus domesticus)[2] 和斑马鱼(Danio rerio)[87] 体内也发现了 γ-HBCD 转化为 α-HBCD 的现象. γ-HBCD 不仅可以转化为α-HBCD,也可以转化为 β-HBCD. Szabo 等[102] 观察到雌鼠 (C57BL/6) 的肝和脑组织中的 γ-HBCD 转变为 β-HBCD,以及脂肪和粪便中的 γ-HBCD 转变为 α-HBCD 和 β-HBCD,该现象也在母鸡 (LohmannBrown)[103] 肝脏中被发现,生物异构化更容易发生在脑、肝脏等具有更长保留时间的组织中.
也有一些研究[104] 发现雌鼠 (C57BL/6) 的脂肪和肝脏中 β-HBCD 可转化为 γ-HBCD.不 仅 γ-HBCD 可 以 转 化 为 α-HBCD, β-HBCD 也 可 以 转 化 为 α-HBCD.Law 等 [89] 发 现 在 虹 鳟 鱼(Oncorhynchus mykiss) 幼体内 β-HBCD 和 γ-HBCD 能异构化为 α-HBCD.同样,在镜鲤 (Cyprimus carpiomorpha moblis)[76] 和蚯蚓 (Metaphire guillelmi、Eisenia fetida)[84, 90] 中,发现了 β-HBCD 和 γ-HBCD 能异构化为 α-HBCD,与 β-HBCD 相比,γ-HBCD 异构化程度更高,且在鲤鱼内脏和肌肉中更容易发生生物异构 化. 该异构化现象也在雄性大鼠( Sprague–Dawley)[88]、小麦( Triticum aestivum L.)[81] 和黑麦草(Lolium perenne L.)[82] 中被发现. Abdallah 等[105] 研究了鼠和鳟鱼(Oncorhynchus mykiss)肝亚细胞组分对 α-HBCD、β-HBCD、γ-HBCD 的体外生物转化,在鳟鱼 S9 馏分(肝匀浆上清液)中检测到 δ-HBCD 说明发生了异构体转化,但具体异构化机制尚不清楚,而鼠 S9 馏分测定中不存在 δ-HBCD,说明鼠体外生物异构化现象与鳟鱼存在明显差异. 一项生物体外转化试验表明鸡肝和猫肝微粒体中 CYP 酶不介导 HBCD 生物异构化和对映选择性代谢[106 − 107].
3.1.2 HBCD 的生物代谢产物HBCD 在生物体内可以通过各种生化反应进行代谢. 相关研究包括一些污染场地生物样品测试和实验室暴露研究.结果表明,HBCD 的生物代谢速度和代谢产物均存在明显的物种特异性[105, 108 − 109],不同异构体的代谢产物亦存在显著差异. 目前关于 HBCD 代谢途径的模型生物主要集中在以小鼠、大鼠为代表的哺乳动物[88, 105, 109 − 111] 和微生物[19, 112 − 118],少量关于禽类、鱼类和植物等[2, 5,76] 的降解研究,以及一些体外实验,如动物肝微粒体[106 − 107] 对 HBCD 的降解.
4 CBFRs 的毒性效应(Toxicity Effect of CBFRs)
4.1 HBCD 的毒性效应HBCD 具有肝脏、神经、免疫、甲状腺、内分泌干扰、生殖发育等毒性效应[8, 125 − 126]. HBCD 是中至强的甲状腺干扰物.HBCD 长期暴露对甲状腺激素三碘甲状腺原氨酸(T3)和四碘甲状腺原氨酸(T4)的分泌与合成有抑制作用[85],能干扰甲状腺激素受体基因(TRβ)的转录[85] 及其代谢酶的活性[127],可能导致甲状腺的肥大[18, 128],此外 HBCD 对促肾上腺激素释放基因(Crh)具有抑制作用[85]. 神经发育毒性是HBCD 的另一个较为敏感的毒性终点.
HBCD 具有抑制神经递质传递[129 − 130]、影响神经信号传导[131]、影响神经元发育[130, 132]、诱导细胞凋亡[130, 133 − 134] 的能力. HBCD 还具有内分泌干扰效应,能通过降低性激素结合球蛋白增加男性游离雄激素指数[135],具有类雌激素效应[136],能与性激素产生拮抗作用[137],影响生殖系统[138 − 139]. 此外 HBCD 在生物体中具有免疫、生长发育、遗传、代谢、肝脏等毒性效应,能够促进脂肪形成基因的表达[140],破坏与免疫和炎症相关的基因表达[18],造成免疫力下降和生长缓慢[141],其引起的代谢功能异常可能导致肥胖症的发展[142].
HBCD 容易在肝脏中积累[143],引起肝脏中 CAR 核受体的活化[18],诱导肝脏重量增加并可能导致肝纤维化[18]. 相关的细胞代谢毒性还能引起 DNA 损伤[117],影响生物体酶的活性[144, 145] 及对多种物质的代谢或运输[18, 105],改变氧化应激关键反应和调控昼夜节律基因的表达[18],影响细胞凋亡和自噬过程[146],引起氧化应激和代谢稳态受损[145],造成代谢网络的破坏[111, 117].此外有研究探讨了 HBCD 对秀丽隐杆线虫(Caenorhabditis elegans)的急性和慢性暴露的毒性效应.在 HBCD 的急性毒性浓度暴露下,最敏感的表现是运动行为的减弱,其次是活性氧和细胞凋亡水平的提高[147],其慢性毒性也包含以上表现[148].大量研究表明,HBCD 的毒性效应亦具有异构体选择性. Palace 等[149] 用喂食的方法将虹鳟鱼分别暴露于 3 种 HBCD 异构体 56 d,其中食物中 α-HBCD、β-HBCD 和 γ-HBCD 的浓度分别为(29.14±1.95)、(11.84±4.62)和(22.84±2.26) ng·g−1 lw,发现不同异构体暴露均可以减少碘的吸收并加速甲状腺激素的转化效率,从而提高 T4 到 T3 的转化率,最终影响虹鳟鱼的代谢,其中 γ-HBCD 的毒害程度最严重.Marteinson 等[150] 发现,以 800 ng·g−1 ww HBCD 的饮食饲养 21 d 后,红隼(Falco sparverius)雏鸟蛋中的HBCD 以 α-HBCD 为主,但羽毛中的以 γ-HBCD 为主. 他们认为羽毛中的 α-HBCD 能够反映 HBCD 的身体负荷,并发现羽毛中的 α-HBCD 浓度与血浆游离三碘甲状腺素与游离甲状腺素的比值呈负相关,能够反应 HBCD 暴露引起的甲状腺功能障碍。
而 β-和 γ-HBCD 含量则没有显著相关性.Du 等[3] 的研究发现,暴露在 0.1、1.0 mg·L−1 水体中,HBCD 的 3 种异构体均能引起斑马鱼(Oncorhynchusmykiss)胚胎发育过程的氧化应激、细胞凋亡、胚胎发育异常,其毒性大小顺序为 γ-HBCD>β-HBCD>αHBCD.但 Hong 等[4] 研究了 HBCD 异构体对海洋青鳉鱼(Oryzias melastigma)胚胎的发育毒性,结果表明在高暴露浓度(200 μg·L−1)下胚胎发育、氧化应激、细胞凋亡,毒性大小顺序为 α-HBCD/γ-HBCD>βHBCD,3种异构体在海水青鳉鱼胚胎的发育毒性顺序与淡水中斑马鱼胚胎中顺序不同,突出了使用淡水和盐水中物种进行毒性评价的重要性. Hong 等[93] 研究了 HBCD 异构体对日本桡足动物(Tigriopusjaponicas)不同发育阶段的毒性,发现不同异构体的急性致死毒性大小为 β-HBCD>α-HBCD>γ-HBCD;在成足期的发育延迟毒性表现为 α-HBCD/γ-HBCD>β-HBCD,且 F1 代对所有 3 种异构体都比 F0 代更敏感.在 30 或 100 μg·L−1 的 α-HBCD、β-HBCD、γ-HBCD 暴露下,产生氧化应激、细胞凋亡、内分泌干扰等毒性效应,其大小顺序为 α-HBCD/β-HBCD>γ-HBCD,可见在同一生物体中急性致死毒性与发育毒性存在异构体特异性.
5 总结与展望(Research Outlook)
CBFRs 可以在生产、使用、运输及废弃物报废处置的整个生命周期中进入大气、水体以及土壤等环境介质,并通过空气和水的流动以及生物的携带进行传输. 近年来,随着 HBCD 生产和使用的限制,预计会有更多的 TBECH 和 TBCO 作为其替代品进入环境,导致他们环境水平的增加. 本文综述了CBFRs 的环境暴露水平、生物富集、转化及毒性效应,重点介绍了 CBFRs 异构体在不同环境介质中的分布,在生物中的富集、代谢及毒性效应的差异性. 针对目前研究的现状以及存在的问题,提出以下总结和展望:
(1)与大量的 HBCD 在环境介质中的时空分布、在环境生物中的富集、代谢、食物链放大和毒性效应研究相比,有关 TBECH 和 TBCO 的研究主要集中于在大气颗粒物及水体沉积物中的分布及在水生生物中的富集和毒性效应,其在陆生生态系统中的研究相当匮乏.为了综合评价 TBECH、TBCO 作为替代品的风险,开展二者的在陆生生态系统的环境行为和生物效应的研究势在必行.
(2)作为 HBCD 潜在的替代品,应该具有更低的生物富集、食物链放大和和毒性效应.目前有关CBFRs 的相关研究各自独立,缺乏与 HBCD 在同样条件下的直接比较,难以对 TBECH 和 TBCO 是否适合做 HBCD 的替代物做出正确评价.因此需要在同一污染场地对三者的生物富集、放大和毒性效应同时进行监测,或在在实验室相同暴露条件的情况下对三者进行研究,同时考察不同种类的生物富集、放大和毒性的差异.(3)TBECH 与 TBCO 具有多种异构体和对映体,异构体和对映体是否存在选择性的生物吸收、是否像 HBCD 一样存在生物异构化反应,异构体及对映体代谢和毒性效应研究均不够全面. 因此,在异构体和对映体水平对以上问题进行研究,是未来工作的重要方向.
( 4) 污 染 物 的 生 物 转 化 是 影 响 污 染 物 的 归 趋 及 毒 性 效 应 的 重 要 因 素.虽 然 目 前 研 究 表 明 ,TBECH 与 TBCO 有较快的生物转化和排出速率,但有限的转化产物的鉴定仅限于体外研究,缺乏体内数据的支持以及对转化途径、转化机制的探索,更缺少在异构体和对映体层面的探索. 因此,有必要开展 TBECH 与 TBCO 在多种生物物种中的体内转化研究,探索转化速率与它们结构的关系. 由于代谢物质缺乏相应的标准品,如何对它们进行定性分析和定量检测,是未来研究亟待解决的关键问题.
(5)CBFRs 在人体内均有检出,具有肝脏、神经、免疫、甲状腺、内分泌干扰、生殖发育等毒性效应. 但目前研究只关注了 CBFRs 原型的毒性效应,它们的一些代谢产物可能呈现比原型更大的生物毒性. 因此,未来在加强 CBFRs 人体暴露风险评估的同时,开展 CBFRs 转化产物的毒性效应研究十分必要.
参考文献 (References)
陈海, 孙云娜, 魏东洋. 溴代阻燃剂的环境危害 [J]. 广东化工, 2011, 38(8): 91-92.CHEN H, SUN Y N, WEI D Y. Bromination flame retardants environmental damage [J]. Guangdong Chemical Industry, 2011,38(8): 91-92(in Chinese).
[ 1 ]FOURNIER A, FEIDT C, MARCHAND P, et al. Kinetic study of γ-hexabromocyclododecane orally given to laying hens (Gallusdomesticus) [J]. Environmental Science and Pollution Research, 2012, 19(2): 440-447.
[ 2 ]DU M M, ZHANG D D, YAN C Z, et al. Developmental toxicity evaluation of three hexabromocyclododecane diastereoisomers onzebrafish embryos [J]. Aquatic Toxicology, 2012, 112/113: 1-10.
[ 3 ]HONG H Z, SHEN R, LIU W X, et al. Developmental toxicity of three hexabromocyclododecane diastereoisomers in embryos of themarine medaka Oryzias melastigma [J]. Marine Pollution Bulletin, 2015, 101(1): 110-118.
[ 4 ]HUANG H L, WANG D, WAN W N, et al. Hexabromocyclododecanes in soils and plants from a plastic waste treatment area in NorthChina: Occurrence, diastereomer- and enantiomer-specific profiles, and metabolization [J]. Environmental Science and PollutionResearch, 2017, 24(27): 21625-21635.
作者:张贞莹1,2 温 蓓1 ** 黄红林1 张淑贞1


Copyright 2002-2023 www.qikanzj.com 京ICP备16051962号