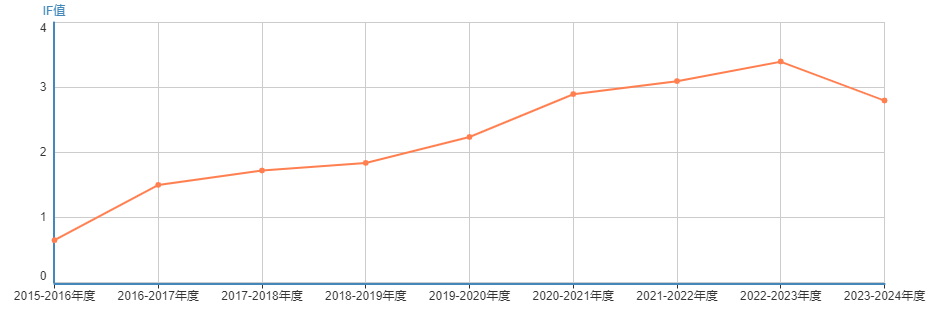
所属栏目:农业论文 时间:2022-05-22
摘 要:尿素是农业生产中施用量最多的氮肥之一,也是植物体内重要的代谢中间产物。普遍认为,植物主要吸收土壤中经土壤微生物转化产生铵态氮和硝态氮,直接吸收尿素和内部水解作用并不显著, 但这一观点正受到挑战,因为植物拥有专用的尿素转运蛋白,能够利用尿素作为唯一氮源。本文通过概述尿素在不同生命系统中存在的基础生理意义及 MIPs 和 DUR3 系统在调控尿素吸收、转运以及库/源间氮循环过程的作用,讨论了脲酶在促进尿素氮的同化、信号传导,精氨酸途径和酰胺途径在维持 C/N 代谢平衡和激素信号网络调节机制,以及植物尿素代谢及稳态调节机制在农业运用方面的展望。通过阐述植物体内局部尿素浓度与整体的氮素状态信号的反馈调节调控氮素的吸收和再利用的复杂调控网络,为提高作物氮的吸收、利用及再分配和农作物遗传改良奠定理论基础。
关键词:尿素;DUR3 蛋白;MIPs 蛋白;脲酶;精氨酸;酰脲

作物的生长发育依赖于土壤中多种营养元素的吸收,尤其受大量营养元素氮(N)供给制约,供应不足通常导致产量和品质下降,严重时导致约50%的产量损失[1]。因此,农业生产中大量增加施氮量以提高作物生产力,减轻日益严峻的粮食安全压力。与硝态氮肥和铵态氮肥相比,尿素由于价格便宜 (低 15% ~ 20%)、含氮量高 (占比 46%)、水溶性高和化学性质相对稳定,以及能与大多数肥料一起混合施用等特性,在多种作物生产中被广泛运用。目前尿素在农业生产中施用量占世界氮肥总消费量的 55%,每年仍以 1.5%的速度递增[2]。农业生产中为追求高产而大量使用化学合成肥料,虽然带来了巨大的经济效益,但是很少有作物可以短期内直接大量利用尿素,过量或不当应用导致氮肥利用效率降低,从而造成生态系统功能的破坏和增加人类健康风险[2-3]。
以中国为例,2017 年使用的尿素为 2.22×107 t,其中有 1.48×107 t 尿素在根际微生物硝化、反硝化作用或土壤脲酶的作用下流失,损失率高达 66.66%,直接经济损失超过 500 亿元人民币(国家统计局,2017. http:// www. stats. gov. cn)。此外,大量施用尿素后,土壤中尿素浓度由通常小于10 μmol·L-1 升高到近 70 mmol·L-1,导致根系大量转运和储存尿素氮,在脲酶作用下会使细胞内氨基酸的浓度增加,特别是谷氨酰胺和天冬酰胺的浓度增加,总氨基酸浓度升高会激活氮素平衡机制,从而抑制植物对尿素的吸收,严重时引起烧苗现象[4-5]。因此,现代农业需要以优化土壤氮和施用氮的平衡,使氮肥施用更具成本性和生态效益兼容性。
目前针对提高作物对尿素利用效率的研究,主要通过根据不同农业生态环境中各栽培品种的需氮特性,制定诸如 4Rs(正确来源、正确比率、正确时间和正确地点)养分管理策略;根据植物-土壤系统中氮代谢途径的生物学、化学和物理学过程,设计新的、更有效的缓控释氮肥材料,以及利用遗传或生物技术方法培育新品种来实现[6]。而对尿素的吸收和代谢研究主要停留在其作为一般氮肥的形式,缺少营养生理学和蛋白质组学等研究来阐明尿素吸收、转运的稳态和信号调控机制。本文综述了尿素 DUR3、MIPs 转运和脲酶参与的尿素吸收和同化机制,以及精氨酸和酰胺参与的氮素循环调控网络的最新研究进展,解析了尿素作用于植物局部或系统水平的生理、形态和分子水平上调节机制,以期从理论上认识与尿素相关的植物氮素营养,为探索提高作物尿素态氮的利用效率开辟新的思路或途径。
1 尿素吸收转运机制对氮素利用的影响
作物尿素的吸收及其积累取决于田间管理、农业实践、环境、遗传特性和其他养分的有效性等,其中氮肥施用和作物遗传特性是影响尿素氮在不同植物中积累的关键因素。由于尿素的分子量较小且其水溶液呈电中性,因此,长期以来尿素分子被认为与水分子类似,易通过自由扩散的方式跨生物膜进入细胞。然而利用 C14 尿素吸收测定法和电生理膜片钳测试技术,发现植物细胞中存在逆电化学梯度的主动吸收途径和膜蛋白促进型的跨膜扩散的被动运输系统[7]。
1.1 尿素高亲和转运的主动运
输曹凤秋等[8]通过与啤酒酵母尿素转运蛋白鉴定对比,发现拟南芥中 DUR3 在尿素浓度极低的条件下也能与驱动二级活性转运的质子一起协同转运尿素,且对尿素的吸收具有专一选择性,表观亲和力常数为 3~4 μmol·L-1,属于高亲和力转运体。AtDUR3主要在根表皮细胞的质膜上表达,转录水平受植物缺氮程度的增加而升高,在缺氮条件下,该蛋白的转录水平更为显著[8]。利用反向遗传法对水稻 dur3突变体在以尿素为单一氮素条件下培养,与野生型相比,OsDUR3 的大量表达使水稻能够在低尿素条件下更好地生长,并吸收更多的尿素,而敲除OsDUR3则导致水稻dur3突变体的根系显著减少且尿素吸收受到抑制[9]。
DUR3 不仅可以增强植物根系对外界环境尿素的吸收,而且在木质部和幼芽附近的质膜上表达,为种子萌发或氮胁迫条件下作为响应因子缓解缺氮影响,参与植物氮素循环中尿素的再分配。例如,水稻种子在萌发过程中,OsDUR3的转录在种子萌发后 1 ~ 6 d 显著增加,并且与胚乳中尿素浓度成正比[10]。在自然或缺氮诱导的衰老过程中,DUR3 基因在维管组织中高度表达,将叶肉细胞中含氮化合物降解产生的尿素回收并转运到线粒体中[11]。在 OsDUR3 缺失突变体 (Tos17)中,尿素因无法正常转运导致在叶片中积累,使圆锥花序不能充分发育,籽粒灌浆量下降近 50%[9]。对拟南芥衰老叶片的叶柄分泌物分析显示,降解产生的尿素占含氮有机物的 13%以上[12]。
1.2 尿素低亲和转运的主动运输
尿素的被动运输主要是由内在蛋白(MIPs)介导的,也称为水通道蛋白,这些蛋白质通过通道沿浓度梯度传导特定的低分子溶质。植物中存在 5 个MIPs (major intrinsic proteins, MIPs)亚科:质膜内在蛋白 (plasma membrane intrinsic proteins, PIPs)、液泡膜内在蛋白 (tonoplast intrinsic proteins, TIPs)、类根瘤-26 的内在蛋白(nodulin 26-like membrane intrinsicproteins, NIPs)、碱性小分子内在蛋白(small, basicmembrane intrinsic proteins, SIPs),以及未知内在蛋白(X-intrinsic Proteins, XIPs),统称为 MIPs [13-14]。MIPs对尿素吸收的动力曲线呈直线型,最高浓度为 30mmol·L-1,这是典型低亲和力转运蛋白的运输特性[15]。
利用绿色荧光蛋白(green fluorescence protein,GFP)进行标记,发现拟南芥 TIPs (AtTIP1.1、AtTIP1.2、AtTIP2.1 和 AtTIP4.1)蛋白在整个植物体中均有较高转录的水平,在 pH 为 5 ~ 8 范围的尿素溶液中活性显著,并且在根系质膜上的转录调控机制与DUR3相似,即受植物组织氮营养状况的调节[16]。此外,TIPs 家族的 TIP1.5 蛋白在液泡膜的位置上,有助于提高拟南芥花粉管中尿素分子从液泡向线粒体基质渗透的效率[17]。这些研究表明 TIPs 蛋白可能既承担了对尿素进行跨质膜运输/吸收的功能,又担负了将细胞质中的尿素转移并贮存于液泡中的职责,从而防止了高浓度尿素在胞质中产生毒害作用,以保证植物的正常发育[18]。PIPs 水选择性孔相对较窄,只允许尿素缓慢通过,与低氮供应相比,高氮供应条件下OsPIP1s 基因表达下调,与此同时叶绿体中的 CO2运输减少,表明 OsPIP1s 基因表达受到组织内 C/N平衡影响[19]。
此外,AtPIP2.1 基因参与叶片生理性缺水诱导的脱落酸形成和调节气孔开关,通过叶片的水分传输影响尿素吸收[20]。而 TIPs 和 NIPs 恰与之相反,可转运足够的尿素,以支持 dur3 缺陷型突变体在仅以尿素为氮源的条件下生长[21]。Grondin等 [22]指出 PIPs 和 TIPs 在植物尿素氮吸收、水分运输和侧根的发育中起着重要的作用,PIPs 在根系吸水贡献率高达 85%。NIPs 的调节具有底物特异性,通过其对水、尿素和甘油,以及硅,硼,锑等金属离子等小溶质的渗透性验证表明,水溶性越好转运效率高[23]。例如,黄瓜在缺氮时可诱导 CsNIP2.1基因的表达,并在以尿素为单一氮源的培养基上使dur3-3 突变体的缺氮症状消失[15]。
相反,在只有尿素供应的培养基中,拟南芥的 AtNIP5.1 和 AtNIP6.1基因表达相对较低,但是相似条件下植物中 DUR3的表达较高,表明 AtNIP5.1 和 AtNIP6.1 的下调与过量尿素水平下尿素/氨的脱毒有关[24]。因此,DUR3和 MIPs 对植物高效利用尿素及维持植物体内的氮营养平衡有重要意义。尿素既可产生于多种氮代谢途径,也能通过位于质膜上的高(HAT,如 DUR3 途径)、低(LAT,如MIPs)亲和力运输系统从环境中进入细胞内。植物中合成尿素的酶主要包括: (1)精氨酸酶;(2)胍丁胺酶;(3)刀豆氨酸水解酶;(4)脒基/酰胺水解酶类,如尿囊酸脒基水解酶和尿素裂解酶。尿素可能通过液泡膜上的转运蛋白(如液泡膜内在蛋白, 即 TIPs)在液泡和细胞质间被区间化。胞质中的尿素经脲酶水解为氨,进而在谷氨酰胺合成酶的作用下进入氮同化途径。
2 脲酶活性对氮同化的调节作用
尿素是否可被植物迅速地利用,取决于植物内外环境中存在的脲酶活性。在植物细胞中,已知能分解尿素的酶有 2 类:脲酶(或尿素酰胺水解酶)和尿素酰胺酶。二者均在细胞质中将尿素水解为 CO2和 NH3,NH3/NH4+随即被谷氨酸合成酶利用,以供其他含氮大分子(蛋白质及核酸等)的生物合成[25]。但是,在高等植物中尿素仅能通过脲酶分解而进入氮同化途径。除从刀豆中提取的刀豆毒素 CNTX(canatoxin, CNTX)是由一个 Ni2+和一个 Zn2+催化外,几乎所有高等植物脲酶都是酰胺水解酶和磷酸三酯酶家族的成员,它们在其活性位点上具有 2 个 Ni2+作为酶活性的活性催化位点[26]。脲酶经过 UreD、UreF、UreG 和 UreE 等辅助蛋白与由赖氨酸羧基化的活性位点,及 4 个组氨酸和一个天冬氨酸残基结合形成羧基桥接两个 Ni2+,后与其他 3 个末端水分子在活性位点形成氢键键合的水四面体簇,从而形成具有稳定催化过渡态产物并加速尿素水解反应的脲酶活性[27]。
Ni2+缺乏极大的降低了脲酶活性,导致尿素在大豆叶片中积累从而影响植物氮素代谢,诱发缺素症状,严重时影响酰脲代谢、尿素循环和尿素吸收[28]。将土壤中 Ni2+浓度从 0.1 mg·kg-1增加到 10mg·kg-1,脲酶活性增加了 20% ~ 30%,大豆芽和叶中氮积累则分别增加了 42%和 50%[29]。通过慢生型大豆根瘤菌(Bradyrhizobium japonicum)评估脲酶在大豆根瘤中的作用发现,大豆和刀豆的脲酶可作为体外细菌细胞识别的趋化因子,促进豆科植物的根瘤固氮作用。与野生型植物相比,缺乏脲酶的突变大豆植株的根瘤较大,但数量较少,这表明脲酶除了参与尿素代谢外,还具有促进尿素吸收的作用[30]。在一些豆科和葫芦科植物的高活性种子中存在一些特异性脲酶:ubSBU (ubiquitous urease, ubSBU)、eSBU (Embryo-specific urease, eSBU)和 SBU-III (enzymatically inactive urease isoform, SBU-III),在参与植物防御机制和促进尿素转运过程中发挥作用[31]。
ubSBU 由 Eu4 基因编码并普遍存在于植物组织中以低水平表达,负责调控内源性或外源性尿素再循环[32]。eSBU 由 Eu1 基因编码,在成熟种子的活性远高于其他植物组织中的 ubSBU 活性,主要参与种子胚胎中氮素积累和种子萌发中氮素代谢[33]。SBU-III是新发现的一种酶促失活的脲酶同种型蛋白,由 Eu5基因编码,在发芽种子、根发育的第一阶段和胚胎发育过程中均有表达,其转录水平低于其他两种大豆同工型脲酶,且 ubSBU 和 eSBU 酶活性较高时,SBU-III 主要参与抵御生物胁迫[34]。通过对大豆幼苗进行人为机械损伤处理后的根和叶片组织采用RT-PCR,对 ubSBU 和 UreG 基因的定量转录水平进行检测,并与脲酶活性进行比较。结果表明 ABA、SA、JA 和 GA 均大量表达并诱导脲酶基因的表达,但脲酶活性不受影响[35]。除此之外,脲酶表达通常受氮代谢的调控,当细胞在 NH4+-N 条件下生长时,酶的合成被阻止,然而尿素或其他氮源存在时,脲酶活性被激活[36]。因此,植物体内脲酶活性是尿素同化效率的重要指征。
3 尿素代谢在氮素积累中的作用
植物在生长过程中需要大量的氮素,尤其是在几个关键的生长节点,环境中有限的氮供应不足以满足植物正常生长需求,植物需要从生物大分子(蛋白质和核酸)中回收和重新分配储存的氮,以满足幼嫩器官的优先发育。例如,氮供应不足或逆境条件下,通过代谢储藏在老叶中的精氨酸或酰胺,可以有效提高叶片水势、光合效率和抗氧化酶活性,并促进渗透压物质的积累以维持内源光合和代谢稳定,以及器官之间的尿素有效转运,从而提高氮素利用率和消除其对环境的负面影响[37-38]。一些证据表明,尿素循环或酰胺代谢循环的激活,也会促进植物激素的表达上升,参与根系发育和氮素调节的长距离信号传导[5]。
4 植物尿素代谢及稳态调节机制
在提高氮素利用率上的展望与铵态氮(NH4+-N)和硝态氮(NO3--N)等化学合成肥料相比,施用于土壤的尿素会被土壤中微生物产生的胞外脲酶快速水解,产生 NH3 挥发到大气中或进一步硝化成 NO3-流失,其水解速率是正常降解速率的 1 014 倍[54, 67]。据估计,全球尿素通过NH3途径平均损失 13% ~ 56%,特别是在轻质土壤、高温高湿和高氮比环境中,NH3 挥发最为严重[68]。
为了解决这些问题,人们开始探索可持续的作物种植方式,以提高氮素利用率。例如,尿素施用后立即灌溉约 15 mm 的水足以将肥料掺入土壤并减少90%的 NH3损失;通过机械或灌溉将尿素掺入土壤,或尿素的掺入深度大于 7.5 cm 时,NH3的挥发可忽略不计[69-70]。基于氮素管理实践,通过添加含硫尿素涂层、有机聚合物涂层、无机-生物涂层、基于尿素作用方式涂层和基于多种营养物质的定向合成涂层制成缓/控释包膜尿素,因其涉及抑制胞外脲酶活性从而降低 NH4+产生和通过硝化抑制剂实现 NH4+硝化至 NO3-的速率减缓,减少系统中尿素氮的损失。利用脲酶是植物从环境中吸收尿素和代谢产生的尿素中的氮再循环所必需,在氮需求旺盛或响应胁迫阶段,植物通过调节尿素代谢或转运机制,刺激氮素调控的网络机制以协调植物尿素的分配途径,使尿素氮转运或分配更有效,从而提高植物氮利用效率[34]。
如在土壤中施用尿素肥料之前,适当的在叶片中喷洒尿素可以提高尿素的吸收效率。根据植物根系对养分的趋向性和地上器官养分信息的系统整合特性,通过一侧施用尿素,而另一侧根系处于氮缺乏状态,建立依赖于外部氮素供应和内源氨基酸库的大小或组成所反映的器官氮状态信号的根系构型(root system architecture, RSA),该过程在氮素缺乏的根系比例不太高(小于 50% ~ 75%),其他的根系就可以在短时间内完全弥补缺氮带来的生长抑制[71-72]。5 结论尿素理化性质比较稳定,其吸收和循环利用过程受土壤尿素浓度和植物氮感受系统的信号网络调节系统调控。
植物种植论文范例:植物保护与病虫害综合防治技术
首先,植物根系对尿素吸收系统 MIPs和 DUR3 转运机制为尿素转运调控的关键转运蛋白的改良和促进根获取尿素氮的灵活性奠定了基础;其次,脲酶在促进尿素氮的利用和信号传导以及在尿素代谢和胁迫响应过程中的作用尤为明显;最后,尿素内源性代谢中,精氨酸途径和酰胺途径对植物生长过程中氮素补充和再利用,以及在种子萌发、生殖生长和响应胁迫过程中维持 C/N 代谢平衡和激素信号传导具有重要的生理意义。然而,在农业生产中尿素的直接利用还有许多的工作要做。关于根吸收系统的结构和调控机制了解不够全面,在模式植物中 DUR3 和 MIPs 转运蛋白家族的功能是否适用与农业生产。在高养分环境下的多年育种栽培,使大部分农作物在养分限制条件下对土壤中N资源的感应和精确定位减弱,也阻碍了关键转运蛋白基因优化育种或生物技术应用。因此,对植物氮素利用过程中氮素感测系统的进一步了解,为探索提高作物尿素态氮的利用效率开辟新的育种思路或途径,提高植物尿素的吸收和代谢具有减少农业系统氮素损失的潜力。例如,可以通过尿素转运蛋白(诱导型)的过表达来增加植物中尿素的摄取或分配。
参考文献:
[1] QBAL J, MITCHELL D C, BARKER D W, et al. Doesnitrogen fertilizer application rate to corn affect nitrousoxide emissions from the rotated soybean crop? [J]. J Environ Qual, 2015, 44(3): 711-719.
[2] DE NOTARIS C, RASMUSSEN J, SØRENSEN P, et al.Nitrogen leaching: a crop rotation perspective on the effect of N surplus, field management and use of catchcrops[J]. Agric Ecosyst Environ, 2018, 255: 1-11.
[3] SHEN Y W, JIAO S Y, MA Z, et al. Humic acid-modifiedbentonite composite material enhances urea-nitrogen useefficiency[J]. Chemosphere, 2020, 255: 126976.
[4] WITTE C P. Urea metabolism in plants[J]. Plant Sci, 2011,180(3): 431-438.
作者:张海涛,宋远辉,郑思怡,花 芹,杨 晔,林泉祥



Copyright 2002-2023 www.qikanzj.com 京ICP备16051962号